






An Enduring Mystery
ARTICLE SUMMARY
• In recent years, controversy has erupted over whether vitamin D2 or vitamin D3 is the predominant form of vitamin D in cod liver oil.
• Research in the 1930s suggested that there were at least four if not six forms of vitamin D in cod liver oil.
• Recent research has shown that fish metabolize vitamin D into at least three other compounds and probably more.
• Although cod liver oil probably does not contain vitamin D2, it probably does contain an array of different compounds derived from vitamin D3.
• The diverse array of vitamin D compounds we would expect to exist in natural cod liver oil likely provides a diverse array of biological activities; many people may experience vitamin D-related benefits from a natural cod liver oil without experiencing as pronounced a rise of 25(OH)D – the blood marker of vitamin D nutritional status—as they would have expected.
• Although it makes sense for someone to increase their sun exposure and vitamin D3 intake if their 25(OH)D is low, low 25(OH)D in and of itself should not be used as evidence that cod liver oil is not providing a vitamin D benefit.
In my 2006 article, “From Seafood to Sunshine: A New Understanding of Vitamin D Safety” (Wise Traditions, Fall 2006), I took the position that vitamin D primarily occurs in cod liver oil as vitamin D3. Nearly a decade later, there are reasons to revisit this question: first, several lab analyses of Green Pasture fermented cod liver oil have come to conflicting conclusions about whether the primary form of vitamin D therein is vitamin D2 or vitamin D3; second, modern research has begun to characterize the way in which fish metabolize vitamin D and has shown that fish contain several different vitamin D compounds. When taken together, the data provide hints that natural cod liver oil may contain a complex array of different vitamin D compounds. The exact nature of vitamin D in cod liver oil, however, remains a mystery in need of resolution.
NATURAL VITAMIN D IS COMPLEX
In an age of fortified foods and vitamin supplements, it is easy for us to overlook the natural complexity of the nutrition found in natural foods. In foods that are fortified with vitamin D, the vitamin D primarily exists in whatever form was added, whether vitamin D2 or vitamin D3. This is not true of natural exposure to vitamin D, whether that exposure comes from the sun or from natural foods.
When sunlight strikes our skin, it converts a close relative of cholesterol into vitamin D. At first, the vitamin D concentration within our skin begins to rise. As vitamin D accumulates, however, the sunlight begins to degrade it into a variety of other compounds. Although researchers generally consider these changes as “degradation” or “inactivation,” those other compounds are not without biological activity: several of them have been shown to prevent the excessive proliferation of skin cells and are thought to help prevent psoriasis.1
When we absorb vitamin D from sun exposure or from our diet, we metabolize it into a plethora of other compounds known as metabolites. The most well known of these is the two-step activation of vitamin D first to 25(OH)D or calcidiol and then to 1,25(OH)2D or calcitriol. These metabolites are also inactivated to 24,25(OH)2D and to 1,24,25(OH)3D. Each of these inactivated metabolites undergoes further metabolism to other compounds. In addition to this major inactivation pathway, there are also several other minor inactivation pathways. Moreover, many of these metabolites exist as stereoisomers, which can best be thought of as mirror images of one another at the level of their chemical structures.
While we often refer to these processes as “activation” or “inactivation,” this is an oversimplification: the changes of biological activity that occur at each step of metabolism are relative rather than absolute, and are often selective. For example, one study in infants found that between 6-18 percent of the circulating 25(OH)D was present as 3-epi-25(OH)D, a stereoisomer or mirror image of the ordinary form. Presumably, this is converted to 3-epi-1,25(OH)2D, which has been found in a variety of cells. When compared to the ordinary form of calcitriol, the 3-epi form has equal ability to suppress parathyroid hormone, a hormone that contributes to bone resorption, and to suppress the excessive proliferation of skin cells; however this form has less power to raise serum calcium levels.
Thus, the range of vitamin D metabolites within our bodies may carry a diverse array of different biological activities rather than each metabolite representing a gain or loss of a single type of biological activity.
Just as this vitamin D metabolism occurs within our own bodies, so it occurs in the animals
hose meat, milk and eggs we eat for food. For example, there are at least five different vitamin D compounds present in natural milk, with the majority of vitamin D activity coming from 25(OH)D, not from vitamin D.2
THE “MULTIPLE NATURE OF VITAMIN D IN FISH OILS”
In “From Seafood to Sunshine,” I cited personal communication with Bruce Hollis for my statement that “only unconverted vitamin D is found in significant amounts in cod liver oil and most other vitamin D-rich foods.” To the best of my recollection from the phone conversation we had while I was writing that article, this was based on speculation about what seemed plausible rather than published data. I also recall from that time that one vitamin D researcher told me privately that I could probably make a lifetime career out of fully characterizing the nature of the vitamins present in natural cod liver oil. I have since encountered a number of reasons to believe the vitamin D in cod liver oil may represent a complex array of compounds.
Research on this topic dates back to the 1930s, much of which is published in the 1937 paper by Charles Bills and colleagues entitled “The Multiple Nature of the Vitamin D in Fish Oils.”3 Bills and his colleagues had shown early on that vitamins D2 and D3 were equally effective in rats but that chickens were unable to use vitamin D2. They then went on to characterize the chick-to-rat “efficacy ratio” of many different marine oils. The logic of the efficacy ratio was as follows: if the oil contained only D3, it would be equally effective in rats and chicks; if the oil was significantly more effective in one or the other species, this would suggest that some or all of its vitamin D activity came from another compound or several other compounds.
The Bills group implemented a number of experimental controls and analyzed a large enough series of oils to rule out the possibility that differences in efficacy resulted from vitamin D being bound to other substances, from synergism or antagonism between vitamins A and D, or from substances within the oil itself that could affect the bioavailability of vitamin D.
Early work in the Bills laboratory had suggested that cod liver oil was equally potent in rats and chickens. This would be consistent with the premise that cod liver oil contains only vitamin D3 and would be inconsistent with the premise that cod liver oil contains only vitamin D2. But these findings may also mean that cod liver oil contains a mix of vitamin D forms that happened to balance out between rats and chickens. In the 1937 paper, the Bills group analyzed the liver oils of three species of codfish: the pollack and two species of hake. They noted that “liver oils of the pollack and these particular hakes are legally cod liver oil” and that they “contribute materially to the ‘cod liver oil’ of certain localities.” The pollack liver oil was only half as effective in chickens than rats, while the hake liver oil was one-third more effective in chickens than rats.
Altogether, they analyzed the efficacy ratios of twenty-five different species and found considerable variation. They concluded that “two (or more than two) kinds of vitamin D exist in fish oils, the proportions varying in the different oils. It now seems unlikely that any particular fish oil, such as cod liver oil, contains one kind exclusively.”
Soon after this, Hickman and Gray, in collaboration with the Bills group,4 subjected cod liver oil to various temperatures to cause specific fractions of the oil to evaporate separately.5 They found that vitamin D activity evaporated into six different fractions, and concluded that cod liver oil contained two major forms of vitamin D and a total of at least four if not six different forms of vitamin D.
Hickman and Gray noted that they used Norwegian cod liver oil, which was “likely to contain more than one species of fish,” so they did not consider their investigation to yield conclusive information about the nature of vitamin D specifically in the species Gadus morhua, known commonly as Atlantic cod. “Nevertheless,” they wrote, “since a wide range of fractions was available from the distillation of many tons of the Norwegian oil, and since the oil continues to be a medicinal favorite, completion of the investigation seemed justified; it was borne in mind, however, that any complexity found in the vitamin D might arise from the mixture of species.”
While this possibility cannot be ignored, it seems quite unlikely that the Norwegian oil would contain liver oils from six different species, each with its own form of vitamin D. It seems far more likely that any one species would have a mix of different vitamin D forms, even if the proportion between those forms might vary from species to species.
At least as late as the 1960s, textbooks cited the Hickman and Gray experiment as showing that vitamins D2 and D3 were the major forms of vitamin D in cod liver oil.6 Hickman and Gray never claimed this, however, because they never performed any analysis of the chemical structures of the vitamin fractions and therefore could not positively identify any of them. The Bills group had considered it “conceivable” but “not especially probable” that vitamin D2 was the component of some marine oils that led to a lower efficacy in chicks than in rats. To my knowledge, the work of Hickman and Gray was never followed up with a definitive identification of the four to six forms of vitamin D they suggested exist in cod liver oil.
MANY VITAMIN D METABOLITES EXIST IN FISH
Although less is known about the metabolism of fish than humans and other mammals, a 2010 review documented a number of studies showing the existence of 25(OH)D, 24,25(OH)2D, and 1,25(OH)2D in addition to vitamin D itself. In the Atlantic cod, about 10 percent of the total circulating vitamin D compounds exists as 24,25(OH)2D and about 13 percent exists as 25(OH)D; there are traces of 1,25(OH)2D and the remainder is vitamin D. Both the liver and kidney of Atlantic cod possess the enzyme that converts 25(OH)D to 1,25(OH)2D. Since the liver is a site of this conversion, one would expect that cod liver oil may contain both 25(OH)D and the fully active hormone, 1,25(OH)2D.
The authors of this review suggested that these metabolites were the “various kinds of vitamin D” that Bills and his colleagues had known existed in fish oils. Based on research in mammals, they also stressed that “for the understanding of the complexity of the vitamin D endocrine system, it is important to realize that many more metabolites of vitamin D3 than described above may exist in fish and could be of physiological significance.”
DOES VITAMIN D2 EXIST IN COD LIVER OIL?
Scientific papers frequently state that although vitamin D2 is known to exist in large amounts in the diet of fish, it has never been found in the flesh or oil of fish. Various suggestions have been offered to explain this, such as the possibilities that fish do not absorb vitamin D2 from the diet or quickly clear it from their systems once absorbed.7 I have had difficulty locating published research documenting the lack of vitamin D2 in fish, but I have personally corresponded with several researchers who say they have often looked for it and never found it.
FIGURE 1. Diagram of an HPLC System
 The diagram depicts the basic features of an HPLC System. The pumps circulate the solvent, known as the mobile phase, through the entire system into the waste container. An injector moves a defined quantity of sample into the mobile phase and the mobile phase carries the sample into the column. Although not depicted, sample injection is typically automated by a computer-controlled autosampler. Separation of compounds within the sample takes place in the column. The mobile phase carries the separated sample compounds into the detector and then into the waste container. The detector sends information about the compounds to a computer, and the computer produces a chromatogram used to estimate the concentrations of the analytes of interest within the sample. See sidebar for a more detailed explanation. This diagram was originally published in Reference 9.
The diagram depicts the basic features of an HPLC System. The pumps circulate the solvent, known as the mobile phase, through the entire system into the waste container. An injector moves a defined quantity of sample into the mobile phase and the mobile phase carries the sample into the column. Although not depicted, sample injection is typically automated by a computer-controlled autosampler. Separation of compounds within the sample takes place in the column. The mobile phase carries the separated sample compounds into the detector and then into the waste container. The detector sends information about the compounds to a computer, and the computer produces a chromatogram used to estimate the concentrations of the analytes of interest within the sample. See sidebar for a more detailed explanation. This diagram was originally published in Reference 9.
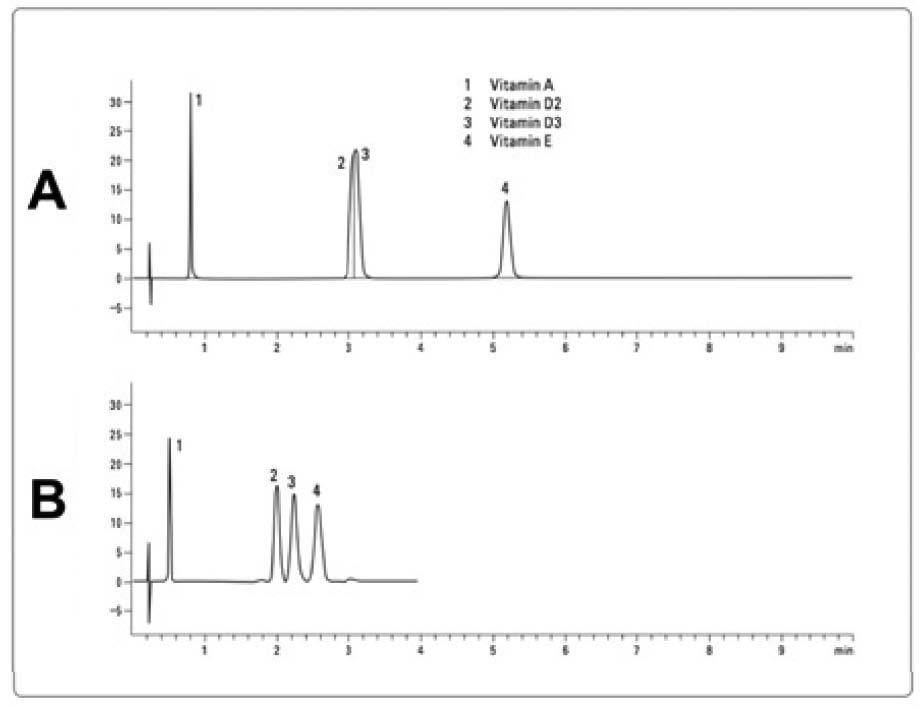 The figure is a slightly edited version of a figure that originally appeared in Reference 11. Both panels of the figure show examples of a chromatogram from an HPLC-UV method. The horizontal axis shows the time in minutes and the vertical axis shows the absorbance of UV light. As compounds that absorb light at the wavelength used elute from the column into the detector, the absorbance suddenly rises and then falls, forming the shape of a peak. The time at which the peak forms is an estimate of the time for which the compound (or compounds) it represents was (or were) retained in the column, and is called the retention time. Comparing panels A and B demonstrates the principle of coelution (see sidebar for a more detailed discussion). In panel A, four vitamins (A, D2, D3, and E) were injected but only three peaks are shown in the chromatogram. Since four peaks were expected, this provided strong evidence that two of the vitamins were coeluting with one another.
The figure is a slightly edited version of a figure that originally appeared in Reference 11. Both panels of the figure show examples of a chromatogram from an HPLC-UV method. The horizontal axis shows the time in minutes and the vertical axis shows the absorbance of UV light. As compounds that absorb light at the wavelength used elute from the column into the detector, the absorbance suddenly rises and then falls, forming the shape of a peak. The time at which the peak forms is an estimate of the time for which the compound (or compounds) it represents was (or were) retained in the column, and is called the retention time. Comparing panels A and B demonstrates the principle of coelution (see sidebar for a more detailed discussion). In panel A, four vitamins (A, D2, D3, and E) were injected but only three peaks are shown in the chromatogram. Since four peaks were expected, this provided strong evidence that two of the vitamins were coeluting with one another.
Although not shown, one could easily deduce which peaks represent which vitamins by injecting only one vitamin at a time and determining its retention time. Numbers have been added to the chromatogram to identify the peaks, and a vertical line has been drawn in the second peak to represent the fact that it reflects two compounds. In panel B, a different column was used that achieved separation between vitamins D2 and D3. Taken together, it is clear that the second peak of panel A is a case of coelution. Nevertheless, the chromatogram in and of itself gives no sign of this: the second peak looks perfectly formed and appears to represent only a single compound. Had this analysis been performed on a complex biological sample such as a natural food instead of a cocktail of four purified vitamins, it would have been much more difficult to determine that the analytes of interest were coeluting.
Vitamin D2 is often added to fish oils as an internal standard (a means of adjusting for loss of vitamin D during sample processing; see sidebar on page 57) when quantifying vitamin D3. Researchers who use vitamin D2 in this manner routinely look for naturally occurring vitamin D2 even if they do not publish the data, because if it exists their use of vitamin D2 as an internal standard would be invalid.
Given this, it was a surprise for two different labs to report that vitamin D2 is the main form of vitamin D in Green Pasture fermented cod liver oil. For a detailed analysis of this topic, see the sidebar on page 63. Readers who are not already familiar with high-performance liquid chromatography (HPLC) and the challenges of using it to measure vitamin D should start by first reading the sidebars on page 57 and 60 to obtain the necessary background information. The conclusion of the analysis in the sidebar on page 63 is that vitamin D2 is probably not present in the fermented cod liver oil.
It seems likely that all natural cod liver oils contain a mix of vitamin D metabolites. These may be exclusively metabolites of vitamin D3, but since the fate of the vitamin D2 known to be present in the diet of fish has not been definitively characterized, the possibility that vitamin D2 metabolites exist in the liver oil should not be ruled out entirely. Additionally, the fermented oil could contain other metabolites of vitamin D produced during the fermentation of the livers.
25(OH)D AS A MARKER OF COD LIVER OIL EFFICACY
In 2009, Green Pasture submitted fermented cod liver oil samples to Deltanoid Pharmaceuticals in Madison, Wisconsin for analysis of biological activity in rats. This analysis concluded that the oil had about 80 IU/mL of vitamin D activity based on the ability of the oil to support normal body weight and normal serum calcium levels.
It is likely that the average human response would be quantitatively different from the average rat response, just as the response from one rat to another or from one human to another would vary. It may also be the case that the oil would better support some biological endpoints than others when compared to other sources of vitamin D. Nevertheless, rats cannot absorb or utilize something from the oil that does not exist, so the rat studies definitively show that the oil is a potent source of one or more compounds responsible for biological vitamin D activity.
At the present time, blood levels of 25(OH) D are almost universally used as the exclusive marker of vitamin D nutritional status. Anecdotally, many people have reported that use of the fermented cod liver oil alone was unable to keep their 25(OH)D levels in the optimal range. It is important to realize that while vitamin D itself will raise 25(OH)D, some vitamin D metabolites would be expected to exert biological activity without raising 25(OH)D. For example, if the fully active 1,25(OH)2D is present in the oil, it will exert most of the biological effects of vitamin D but it will not raise 25(OH)D. In fact, it could even lower 25(OH)D by convincing the body that less 25(OH)D is needed and shunting it into an inactivation pathway.
I think it is wise to diversify one’s vitamin D intake between sun exposure and various dietary sources rather than relying on one product alone. However, I have also come to believe that 25(OH) D is being overused as a marker of vitamin D nutritional status and is being used in an overly simplistic manner. As I have been documenting in my “An Ancestral Perspective on Vitamin D Status” series on the Mother Nature Obeyed blog of westonaprice.org, several populations seem to be genetically adapted to a lower 25(OH)D level than American and European whites and there are many factors—some good, such as vitamin A; others bad, such as calcium deficiency or inflammation—that can lower 25(OH)D besides an inadequate intake of vitamin D.
AN ENDURING MYSTERY
The exact nature of vitamin D in cod liver oil remains to be fully elucidated. It is not a mystery in the sense that it cannot be resolved, but it endures as a mystery in the sense that much remains to be learned. I consider it likely that as we learn more, we will come to a greater and greater appreciation of the magnificent complexity contained within natural foods.
SIDEBARS
AN INTRODUCTION TO HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY (HPLC)8.9
High-performance liquid chromatography (HPLC) is a means of separating individual compounds within a sample. Sometimes, the separation serves to isolate or concentrate a compound or a collection of compounds so that a scientist may use them for other purposes. This is called preparative HPLC. In this article, we are concerned only with HPLC that separates compounds for the purpose of measuring their concentrations within a sample, which is called analytical HPLC. In these cases, the compounds being analyzed are called analytes. In analyses aimed at quantifying the amount of vitamin D2 and vitamin D3 in a sample, we would say that vitamins D2 and D3 are the analytes of interest.
The basic setup of an HPLC system is shown in Figure 1. A pump or set of pumps moves a fluid called the mobile phase from its initial reservoir, typically a glass bottle, through the entire system and into a waste container. The person operating the system may have prepared the samples in any number of ways to make them more compatible with the method, and will have loaded them into the autosampler, a machine that automatically injects a defined quantity of sample into the mobile phase at specific time points determined by a computer program. Once the sample is injected, the mobile phase carries it to the column, where separation takes place. The separated analytes exit the column, one by one, into a detector. This process is called elution and the analytes are said to elute from the column. The detector then collects information that can be used to quantify the compounds and sends that information to a computer. Finally, the person operating the system uses a software application on the computer to analyze the information and thereby infer the concentration of the analytes in the sample.
While the fluid mobile phase carries the sample through the column, the column itself contains a solid bedding called the stationary phase. The principle of separation is competitive attraction between the mobile and stationary phases: all the compounds in the sample will have some level of attraction to the mobile phase and some level of attraction to the stationary phase; each particular compound, however, will have its own balance between these two attractions and will therefore pass through the column at its own rate. Thus, while the injected sample first arrives at the column as a single unit, the compounds that are most attracted to the mobile phase and least attracted to the stationary phase will elute from the column first, and the compounds with the opposite characteristics will elute last.
As the analytes elute from the column, they pass into the detector. There are a variety of detector types: some, for example, detect the ability of the analytes to absorb ultraviolet (UV) light, to absorb visible light, to fluoresce, or to undergo chemical reactions. The detector sends this information to a computer, and the computer software creates an image from it called a chromatogram (Figure 2). The chromatogram shows a series of peaks, each representing material that eluted from the column at a specific time point. That time is called the retention time because it reflects the amount of time for which the material that generated the peak had been retained in the column.
To quantify the analytes of interest in a sample, the magnitudes of their peaks (often measured as the peak area or height) are compared to those generated by known quantities of a standard. The standard can be external or internal. An external standard is a known quantity of the pure analyte of interest prepared in the same way and at the same time as the samples but injected separately into the HPLC system so that it generates its own chromatogram. An internal standard is a compound that is very similar to but distinct from the analyte of interest that is added to each sample at the beginning of the preparation step, injected into the system as part of each sample, and generates its own peak within the chromatogram of each sample. External standards help define how the size of the peaks relates to the concentration of the analyte of interest and help control for variation between batches in that relationship. Internal standards help control for loss of the analyte during sample processing. Depending on the relative need for these controls, a method may call for one, the other, or both.
Readers interested in learning more about chromatography should consult Reference 8, an essential text on the subject. General information on HPLC within this sidebar and throughout this article that is not associated with a specific citation is drawn from this reference as well as from my own laboratory experience.
CHALLENGES USING HPLC TO MEASURE VITAMIN D
Measuring vitamin D with HPLC (see sidebar on page 57 for a general introduction to HPLC) presents numerous challenges that give the several different methods used their own strengths and limitations. These challenges can best be understood with an appreciation of how the different detectors used to measure vitamin D work.
The most common HPLC method for measuring vitamin D utilizes a detector that measures its absorption of UV light. The detector is called a UV detector, and the method is abbreviated as HPLC-UV. Another way to measure vitamin D is with mass spectrometry (MS). The simplest version of this method adds a charge to the analyte; the detector distinguishes between the analyte of interest and other analytes based on their mass-to-charge ratios. The detector is called a single-stage mass spectral detector and the method is abbreviated LC-MS. A more sophisticated version of MS fragments the charged analyte into smaller pieces; each analyte fragments in a characteristic pattern, giving a specific “signature” of mass-to-charge ratios. This type of detection is called tandem MS, its use to monitor the fragmentation pattern of an analyte is called multiple reaction monitoring, and the method is abbreviated LC-MS/MS.
One can appreciate the difference in specificity between these methods by likening them to the specificity of different methods to identify a person. HPLC-UV is like seeing someone—anyone—in a particular place at a particular time. If you know enough about the context—where the person came from, where they were going, who they interacted with —you can reasonably deduce that the person is who you think it is, but you cannot be completely certain. LC-MS is like identifying someone based on their shape, size, and facial characteristics. LC-MS/MS is like identifying someone based on their fingerprints. In the order HPLC-UV, LC-MS, LC-MS/MS, these methods are not only more specific but also more sensitive, meaning they are better able to detect small traces of compounds. But the equipment for the more sensitive and specific methods is also more expensive to purchase, operate, and maintain, and the technical expertise needed to operate the equipment and analyze the results is less common. Which method to use depends on which challenges one faces in the specific analysis.
The primary challenges in measuring vitamin D relate to a concept known as coelution. In a perfect separation, each compound would elute from the column all by itself, and each peak in the chromatogram would represent a singular substance. In practice, however, compounds often elute together. This is called coelution and the compounds are said to coelute. This is desirable if the coeluting compounds are not the analytes of interest: if you could get the analytes of interest to separate perfectly and everything else to coelute, it would take a lot less time to do the analysis. Coelution can also be irrelevant if the coeluting substances are not picked up by the detector. For example, in a common HPLC-UV method, there is a triglyceride in fish oil that coelutes with vitamin D2; since that triglyceride does not absorb UV light, however, it does not contribute to the vitamin D2 peak in the chromatogram and does not interfere with the detection of the vitamin.10
Coelution becomes a major problem when either two or more analytes of interest coelute with one another, or when some other substance that generates a signal in the detector coelutes with an analyte of interest. Sometimes the problem is obvious because the peak of interest looks deformed or because one of the peaks of interest is missing, and often the problem can be fixed simply by modifying the method to achieve better separation. For example, the chromatography company Agilent advertises an HPLC column that generates only three peaks in response to vitamins A, D2, D3, and E (Figure 2A); one of those peaks represents vitamins D2 and D3 coeluting together, but when a different Agilent column is used for the same method, D2 and D3 elute separately and each of the four vitamins generates its own peak (Figure 2B).11
Sometimes, however, the problem is not obvious at all: what looks like a single, well-formed peak could actually represent the analyte of interest jumbled together with one or more unidentified coeluting substances. For example, when a common HPLC-UV method for measuring vitamin D3is used on processed cheese, the D3 peak appears normal but more than two thirds of the peak area reflects unknown coeluting substances.12
In this case, proper use of HPLC-UV could identify the problem by showing the peak to be impure. The peaks in an HPLC-UV chromatogram represent absorption of UV light of a single wavelength; in the case of vitamin D, the wavelength is 265 nanometers (nm). The purity of these peaks can be assessed by conducting a full wavelength scan. Many compounds may absorb UV light at 265 nm, but individual compounds tend to have their own characteristic absorption spectrum, with maximum absorption at one wavelength, minimum absorption at another, and a predictable pattern for the wavelengths in between (Figure 3A). The putative “D3” peak in processed cheese has an absorption pattern that is clearly distinguishable from that of pure D3, making it clear the peak is contaminated.
Although a full wavelength scan can help identify the problem by showing that the peak is impure, it cannot solve the problem by quantifying the proportion of the peak that reflects true vitamin D3. The only way to do this while still using UV detection would be to modify the method in a way that achieves separation of vitamin D3 from the coeluting substances; this is time- and labor-intensive, and many laboratories would not have the resources to solve the problem in this way. Another way to solve the problem is to use LC-MS. In the case of the processed cheese, LC-MS showed that only one-third of the substances that contributed to the “D3” peak in UV analysis had the mass that would be expected for D3; thus, LC-MS generated a much lower and more accurate estimate for the vitamin D3 content of the cheese. Since the compounds that coelute with D3 in this case have not been identified, one could argue that some of them do generate an ion with the same mass as that generated by D3 and that even LC-MS is arriving at an overestimation. If this were suspected, LC-MS/MS could offer even greater specificity.
One area where MS becomes especially important is in distinguishing the many vitamin D compounds from one another.13 Vitamins D2 and D3, as well as most of their metabolites, share the same UV absorption spectrum (Figure 3). If they were to coelute with one another, HPLC-UV would not even be able to generate evidence of peak impurity. HPLCUV also lacks the sensitivity to detect most vitamin D metabolites at the low concentrations in which we would expect them to occur in foods. There are small differences in mass between vitamins D2 and D3, and between the parent vitamins and some of their metabolites, allowing LC-MS to make many distinctions that HPLC-UV cannot. Some metabolites of vitamin D, however, have identical masses, limiting the usefulness of LC-MS in some contexts. LC-MS/MS is ideal for these analyses because of its greater specificity and sensitivity.
Even LC-MS/MS faces challenges.13 One of those has been to distinguish 25(OH)D from 3-epi-25(OH)D. This has been important because both compounds occur in human blood but they do not have the same biological activity. They are stereoisomers of one another, which means that their chemical structures are like mirror images. They generate identical fragmentation signatures, so even an LC-MS/MS detector cannot tell the difference between them. Analytical chemists have therefore been focused on getting the two compounds to elute separately so that they can be distinguished based on their retention times. LC-MS/MS is also usually insufficient for characterizing the structures of previously undiscovered vitamin D-related compounds. This has generally required the combination of mass spectrometry with gas chromatography. Gas chromatography is similar to liquid chromatography except that the solvents and samples move through the machine as gases rather than liquids, and the ionization procedure used in these methods can generate more specific fragmentation signatures.
Thus, HPLC-UV is ideal for routine quantification of nutrients in simple, well characterized samples, while LC-MS or LC-MS/MS is ideal for analyzing nutrients in complex or poorly characterized samples, and gas chromatography is usually a necessary complement to these methods when operating in truly uncharted scientific territory.
DOES THE VITAMIN D ACTIVITY OF FERMENTED COD LIVER OIL COME FROM VITAMIN D2?
As discussed in the main text, tests of biological activity conducted in rats show that the Green Pasture fermented cod liver oil is a potent source of vitamin D activity. The possibility that vitamin D2 may be the primary form of vitamin D in this oil arose when lab results from UBE Analytical Laboratories in California and Nutrasource Diagnostics, Inc. (NDI) in Ontario, Canada reported that this was so. More recently, Dr. Kaayla Daniel submitted the product to two laboratories that found vitamin D3 in the oil but no detectable D2.14
One of the labs that Dr. Daniel used analyzed the oil with LC-MS (not LC-MS/MS). The method used by the other laboratory is unclear based on the data provided in the report. Unfortunately, the data were obtained with non-disclosure agreements that preclude a critical analysis for this article. Dave Wetzel of Green Pasture generously gave me full access to the lab reports from UBE and NDI and allowed me to publish them and critically analyze them in this article. A representative from NDI tried to answer my questions about the laboratory methods but after several days of research into internal records told me that they had been subcontracted to another laboratory in 2007 and it was no longer possible to obtain the detail that would be necessary for this article. Fortunately, Danny Pang, the laboratory manager of UBE, was willing and able to answer all of my questions and I am therefore able to produce a critical analysis of the UBE results herein.
UBE analyzes the oil with HPLC-UV (see the sidebars on pages 57 and 60 for a detailed discussion about the strengths and limitations of this method compared to other methods). As can be seen in Figure 4A, the method efficiently separates pure vitamin D2 and D3 external standards. As can be seen in Figure 4B, there is a single putative vitamin D peak in the fermented cod liver oil, and its retention time is, while not exactly the same as either standard, closer to that of the vitamin D2 standard than to that of the vitamin D3 standard. Pang told me that they called the peak “vitamin D2” based solely on its retention time. Since the UV spectra for vitamins D2 and D3 are identical, retention time is the only tool available to distinguish between the two in an HPLC-UV method. Pang also told me, however, that he suspected the peak may reflect a microbial metabolite of vitamin D produced during the fermentation process and that it may be better to refer to it as a D2-like compound.
One problem with concluding that the peak represents D2 based on the retention time alone is that the retention time of a peak can be affected by the matrix in which the analyte is injected into the HPLC. The standards are dissolved in a chemical solvent, while the vitamin D compounds are dissolved in the fermented cod liver oil itself. I asked Pang whether they had tried spiking the fermented cod liver oil with known, purified D2. If the known D2 coeluted with the putative D2 peak in the oil, this would offer some additional support for the conclusion that the peak truly represents D2. Pang told me that they had not done so yet because of limitations on their time. However, in response to our conversation Pang had the samples reanalyzed with and without being spiked with known vitamin D2. As can be seen in Figure 4C, the known vitamin D2 does not coelute with the putative D2, strongly suggesting that the peak does not represent D2 itself.
At the time of writing, I do not have access to any data showing whether pure D3 coelutes with the unknown peak in the oil. Since the unknown peak in the oil elutes earlier than D2 (Figure 4C), whereas the pure D3 standard elutes later (Figure 4A), this seems very unlikely; however, it is not completely impossible that the sample matrix could affect the order of the peaks. Without conclusively identifying the specific compound(s) responsible for the peak, moreover, it is impossible to know whether it represents one compound or a collection of two or more compounds. It is entirely within the realm of possibility that multiple vitamin D compounds, possibly including metabolites produced by the fish as well as microbial metabolites produced during fermentation, are coeluting together. If such metabolites are present in the oil, fully characterizing its vitamin D activity would require a very expensive, time-intensive, and labor-intensive process using more sophisticated chromatography methods.
Overall, the data suggest that the vitamin D activity of the fermented cod liver oil does not come from vitamin D2, but may not come from vitamin D3 either; at present, the vitamin D activity of the oil should be regarded as mystery in need of further resolution.
FIGURE 3: Most Vitamin D Compounds Share the Same UV Absorption Spectrum
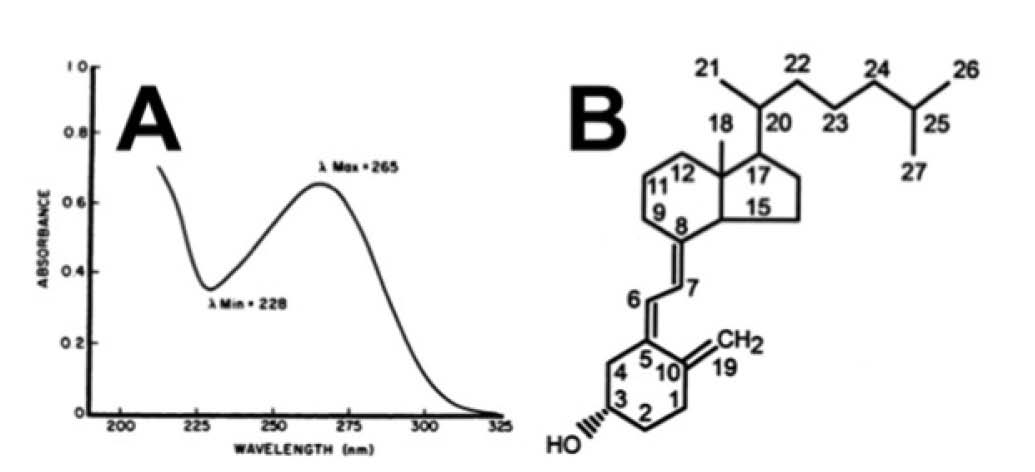 Any compound that absorbs UV light has a characteristic absorption spectrum. Panel A shows the absorption spectrum of vitamin D and was originally published in Reference 15. Wavelength is abbreviated by the Greek letter lamda (l). The wavelength at which vitamin D absorbs the most UV light is known as its l max and is 265 nm. The wavelength at which it absorbs the least is known as its l min and is 228 nm. A putative vitamin D peak in an HPLC chromatogram could be shown to be misidentified or impure if the absorption spectrum clearly differed from that of pure vitamin D. Solvents used in HPLC absorb UV light, and UV spectra often show additional absorbance at lower wavelengths because of the solvent, as is the case in panel A.
Any compound that absorbs UV light has a characteristic absorption spectrum. Panel A shows the absorption spectrum of vitamin D and was originally published in Reference 15. Wavelength is abbreviated by the Greek letter lamda (l). The wavelength at which vitamin D absorbs the most UV light is known as its l max and is 265 nm. The wavelength at which it absorbs the least is known as its l min and is 228 nm. A putative vitamin D peak in an HPLC chromatogram could be shown to be misidentified or impure if the absorption spectrum clearly differed from that of pure vitamin D. Solvents used in HPLC absorb UV light, and UV spectra often show additional absorbance at lower wavelengths because of the solvent, as is the case in panel A.
Although the absorption spectrum of any given compound tends to differ from that of most others, most vitamin D compounds share identical absorption spectra because they share in common the structural feature responsible for UV absorption. Panel B shows the structure of vitamin D with each carbon atom assigned a number. The portion of the molecule that absorbs UV light is the set of three double bonds (symbolized by three sets of double lines and known as a triene structure, “tri-” for three and “-ene” for double bonds) between carbons 5 and 6, carbons 7 and 8, and carbons 10 and 19. Most vitamin D compounds retain this structural feature intact. For example, D2 differs from D3 by having an extra carbon atom attached to carbon 24; the main activation pathway of vitamin D involves hydroxylation of carbons 25 and 1; the main inactivation pathway involves hydroxylation of carbon 24. All of these modifications occur on the outside rings or the side chain of the molecule and leave the UV-absorbing triene structure unchanged.
FIGURE 4: HPLC Analysis of the Fermented Cod Liver Oil
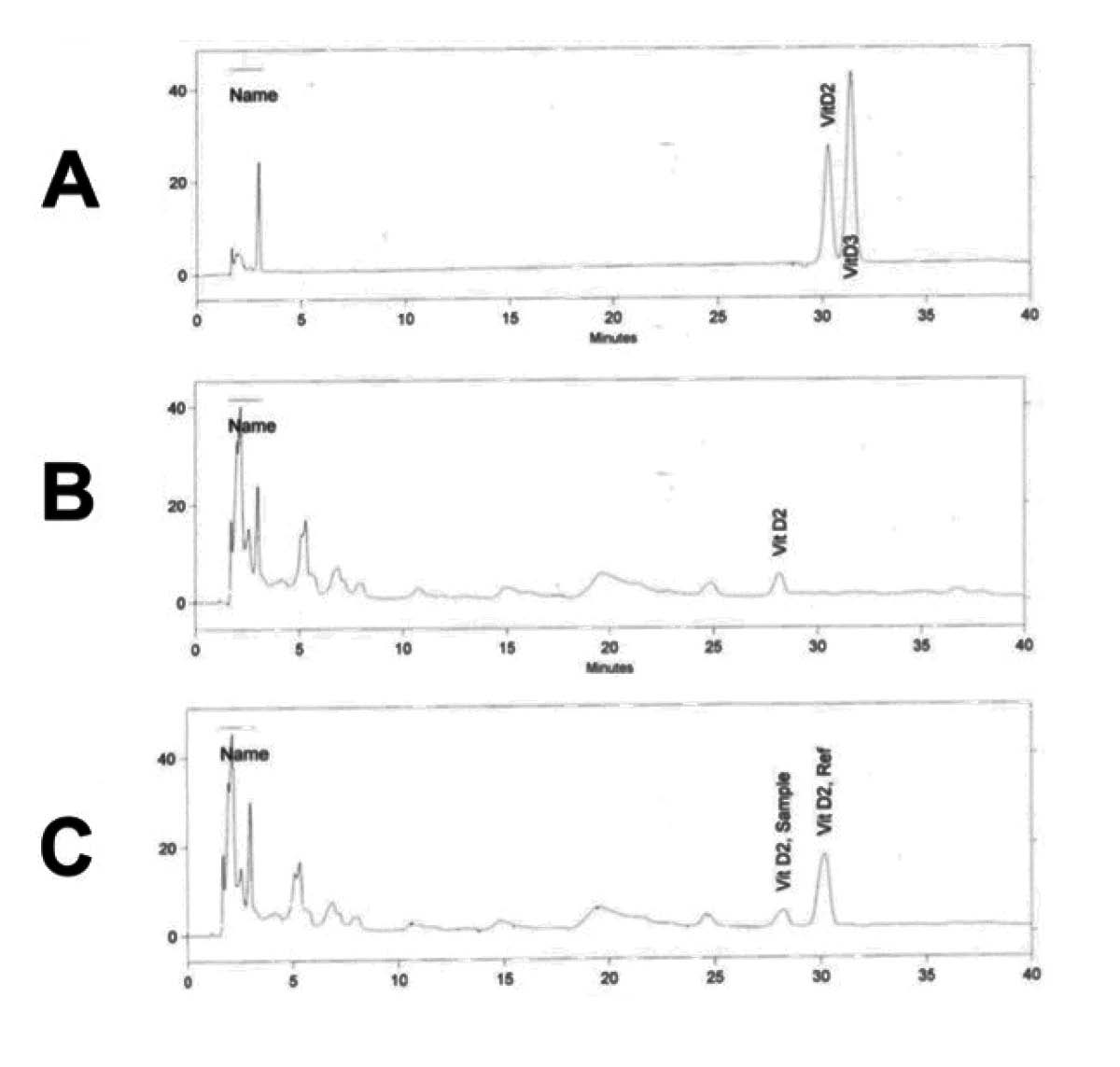 The figure shows HPLC-UV analysis of Green Pasture fermented cod liver oil, conducted by UBE Laboratories. Panel A is a chromatogram of the external standards and shows that the method satisfactorily separates vitamins D2 and D3. Panel B is a chromatogram of the oil. A comparison of panels A and B shows that the putative D2 peak in the oil has a retention time (28.1) closer to that of the D2 standard (30.3) than to that of the D3 standard (31.4). Panel C shows the chromatogram of fermented cod liver oil that has been spiked with known D2. If the putative D2 peak (Vit D2, Sample) truly represents D2, it should coelute with the D2 that had been added (Vit D2, Ref). This does not happen: the putative D2 peak elutes at 28.2 min, while the known D2 elutes at 30.2 min. This suggests that the putative D2 peak does not represent true D2.
The figure shows HPLC-UV analysis of Green Pasture fermented cod liver oil, conducted by UBE Laboratories. Panel A is a chromatogram of the external standards and shows that the method satisfactorily separates vitamins D2 and D3. Panel B is a chromatogram of the oil. A comparison of panels A and B shows that the putative D2 peak in the oil has a retention time (28.1) closer to that of the D2 standard (30.3) than to that of the D3 standard (31.4). Panel C shows the chromatogram of fermented cod liver oil that has been spiked with known D2. If the putative D2 peak (Vit D2, Sample) truly represents D2, it should coelute with the D2 that had been added (Vit D2, Ref). This does not happen: the putative D2 peak elutes at 28.2 min, while the known D2 elutes at 30.2 min. This suggests that the putative D2 peak does not represent true D2.
REFERENCES
1. Chen, T. C. Persons, K. S. Lu, Z. Mathieu, J. S. Holick, M. F. An evaluation of the biologic activity and vitamin D receptor binding affinity of the photoisomers of vitamin D3 and previtamin D3. J. Nutr. Biochem., 2000; 11: 267-272.
2. Hollis, B. W. Roos, B. A. Draper, H. H. Lambert, P. W. “Vitamin D and Its Metabolites in Human and Bovine Milk,” J. Nutr., 1981; 111: 1240-1248.
3. Bills CE, Massengale ON, Imboden M, Hall H. The Mulitple Nature of the Vitamin D of Fish Oils. J Nutr. 1937;13(4):435-52.
4. Bills CE, Massengale ON, Hickman KCD, Gray EL. A New Vitamin D in Cod Liver Oil. J Biol Chem. 1938;126:241-44.
5. Hickman KCD, Gray EL. Molecular Distillation. Examination of Natural Vitamin D. Ind Eng Chem. 1938;30(7):796-802.
6. Higashi H. Vitamins in Fish – with Special Reference to Edible Parts. In Borgstrom G., Ed. Fish as Food V1: Production, Biochemistry and Microbiology. 1961.
7. Lock EJ, Waagbo R, Wendelaar Bonga S, Flik G. The significance of vitamin D for fish: a review. Aquaculture Nutrition. 2010 16;100-116.
8. Snyder LR, Kirkland JJ, Dolan JW. Introduction to Modern Liquid Chromatography: Third Edition. Wiley, 2009.
9. Czaplicki, S. (2013). Chromatography in Bioactivity Analysis of Compounds, Column Chromatography, Dr. Dean Martin (Ed.), ISBN: 978-953-51-1074-3, InTech, DOI: 10.5772/55620. Available from: http://www.intechopen.com/books/column-chromatography/chromatography-in-bioactivity-analysis-of-compounds
10. Byrdwell WC. “Dilute-and-shoot” triple parallel mass spectrometry method for analysis of vitamin D and triacylglycerols in dietary supplements. Anal Bioanal Chem. 2011; 401(10): 3317-34.
11. Ronjie FU. Fat-Soluble Vitamins Analysis on an Agilent ZORBAX Eclipse PAH Polymeric C18 Bonded Column. Agilent Technologies. February 15, 2010. 5990-5342EN.
12. Byrdwell WC. Comparison of Analysis of Vitamin D in Foods Using Ultraviolet and Mass Spectrometric Detection. J. Agric Food Chem. 2009(57(6):2135-46.
13. Gathungu RM, Flarakos CC, Styanarayna Reddy G, Vouros P. The role of mass spectrometry in the analysis of vitamin D compounds. Mass Spectrometry Reviews. 2013;32:72-86.
14. Daniel, KT. Hook, Line, and Stinker: The Truth about Fermented Cod Liver Oil. http://drkaayladaniel.com/hook-line-and-stinker-opt-in/ Published and accessed in August, 2015.
15. Hollis BW, Roos BA, Draper HH, Lambert PW. Occurrence of Vitamin D Sulfate in Human Milk Whey. J Nutr; 1981. J Nutr: 111:384-90.
This article appeared in Wise Traditions in Food, Farming and the Healing Arts, the quarterly journal of the Weston A. Price Foundation, Fall 2015
🖨️ Print post
Chris, my doctor wants me to take supplemental vitamin D. She originally gave me a prescription for a once a month 50,000 i.u. D2 pill. I told her I’d feel better taking the D3 form. However, I have also discovered that I have an strong allergic reaction (intense rash on hands) to anything I put on my body or in my body that is sourced from lanolin. I’ve also discovered that most, if not all, of the high potency, over the counter, D3 supplements are sourced from lanolin. I have found one brand (Solaray) that is 400 iu D3 per gel cap sourced from fish liver oil. So, I take 5 of these gel caps a day. Do you know of other brands that are high potency D3 fish liver oil?
Low d is really low magnesium as you need magnesium to synthesize d. Also they share the same receptor so when you take synthetic d without magnesium you can cause a magnesium deficiency. Best way to get d is from the sun and cod liver oil. Rosita’s is a virgin inprocessed brand you can try.
This completely clears up all the confusion. I will will be sharing on many Youtube videos that are currently bashing this most important super FOOD. Thank you.
Also, to anyone who may be interested, please look into the the unique light filtering technology discovered in Egyptian mines. They recreated/replicated the storage device and it has been named MIRON GLASS. It blocks all Harmful light rays EXCEPT for the violet part. At the same time it allows a certain part to be permeable for radiation in the spectral range of UV-A, and infra red light! This unique combination offers optimal protection against the ageing processes that are released by visible light, thus lengthening durability and potency of products! I store my Fermented Cod Liver in it and definitely notice an improvement in the oil compared to my other amber bottles of FCLO.
God-speed, it means Best of Wishes!
Wow what a great article! I’m just upset with myself for being easily confused about sources of vitamin D. I realized this when I spoke with someone at Nordic Naturals and asked why there is no Vitamin D on the label of their liquid cod liver oil. Couldnt believe what I was hearing actually. So, is the level of Vitamin D on the label of other products mean they are adding D3/D2 to their products? My only concern is that I have contracted COVID three times and the last time I barely pulled through. I’m trying to develop a program to increase my blood serum of D3 and its metabolites.
Thank you so much.
Jose