





 🖨️ Print post
🖨️ Print post
Summary
- Vitamins A, D, and K2 interact synergistically to support immune health, provide for adequate growth, support strong bones and teeth, and protect soft tissues from calcification.
- Magnesium is required for the production of all proteins, including those that interact with vitamins A and D.
- Vitamins A and D support the absorption of zinc and zinc supports the absorption of all the fat-soluble vitamins.
- Many of the proteins involved in vitamin A metabolism and the receptors for both vitamins A and D only function correctly in the presence of zinc.
- Dietary fat is necessary for the absorption of fat-soluble vitamins.
- Vitamin K2 activates proteins by adding carbon dioxide to them. We can increase carbon dioxide production by consuming carbohydrates, exercising, and maintaining robust thyroid status.
- The fat-soluble vitamins thus interact not only with each other, but with other nutrients and metabolic factors as well. This level of complexity should urge us to exercise caution when interpreting scientific research, and to cooperate with the wisdom of nature by obtaining vitamins through nutrient-dense foods.
Throughout the twentieth century, we viewed the fat-soluble vitamins in isolation from one another. Researchers relegated vitamin K to blood coagulation, and ignored vitamin K2 entirely because it is present in such small quantities in the diet. Vitamins A and D each fell in and out of favor, the popularity of one always rising at the expense of the other. Ignoring the forest to study one tree at a time, the concept of synergy eluded us.
In order to truly understand the fat-soluble vitamins, however, we must understand that vitamins A, D, and K cooperate synergistically not only with each other, but also with essential minerals like magnesium and zinc, with dietary fat, and with key metabolic factors like carbon dioxide and thyroid hormone.
Vitamins A, D, and K
The “K” in “vitamin K” stands for “koagulation,” the German word for blood clotting. From its discovery in the 1930s through the late 1970s, we knew of no other roles for the vitamin. The 1990s had come and nearly gone by the time awareness of its role in bone metabolism broke out of the confines of the vitamin K research community, and only in the twenty-first century has its role in preventing calcification of the blood vessels and other soft tissues become clear. Vitamin K2, found in animal fats and fermented foods, is present in much smaller quantities in most diets when compared to vitamin K1, found in leafy greens. Since researchers throughout the twentieth century saw the two forms of the vitamin as interchangeable, they ignored vitamin K2 as though its scarcity made it irrelevant. The realization that vitamin K is not just for “koagulation,” however, led us to discover that vitamins K1 and K2 are not interchangeable after all: vitamin K1 more effectively supports blood clotting, while vitamin K2 more effectively ensures that calcium winds up in the bones and teeth where it supports health rather than in the soft tissues where it contributes to disease. It was thus only in 2006 that the United States Department of Agriculture determined the vitamin K2 contents of common foods for the first time.1
While vitamin K2 languished in obscurity, vitamins A and D continually traded places with one another as the favored vitamin du jour. The pendulum initially swung in favor of vitamin D because rickets was common in the early twentieth century while eye diseases resulting from vitamin A deficiency were rare. It then swung in favor of vitamin A when that vitamin became known as the “anti-infective” vitamin.2 After World War II, the medical establishment had easy access to antibiotics and thus lost interest in battling infections with vitamin A.3 Vitamin D fared far worse, taking the blame for a British epidemic of infant hypercalcemia and eventually earning a reputation as “the most toxic of all the vitamins.”4
These days, the pendulum has swung full force in the opposite direction: we blame an epidemic of osteoporosis on vitamin A, and see vitamin D as the new panacea.5
Though a paradigm of synergy never took hold, it was not for want of opportunity. When Mellanby and Green first demonstrated in the 1920s that vitamin A prevented infections, they concluded that vitamin D could not be “safely substituted for cod-liver oil in medical treatment,” and that “if a substitute for cod-liver oil is given it ought to be at least as powerful as this oil in its content of both vitamins A and D.”2 Consistent with this point of view, clinical trials in the 1930s showed that cod liver oil could reduce the incidence of colds by a third and cut hours missed from work in half.6 Cod liver oil also caused dramatic reductions in mortality from less common but more severe infections. The medical establishment, for example, had been successfully using it to treat tuberculosis since the mid-nineteenth century.7 Studies in the 1930s expanded this to the treatment of measles.8 These findings made the popularity of cod liver oil soar (Figure 1).9
The idea that vitamin A alone was “antiinfective,” however, led to similar trials with halibut liver oil, which is rich in vitamin A but poor in vitamin D. These trials often failed to show any benefit. I.G. Spiesman of the University of Illinois College of Medicine proposed a simple solution to this paradox: vitamins A and D worked together to prevent infection, he suggested, and both vitamins are needed to prevent the common cold. He published his own clinical trial in 1941, showing that massive doses of each vitamin alone provided no benefit and often proved toxic. Massive doses of both vitamins together, however, caused no toxicity and offered powerful protection against the common cold.10 Nevertheless, as antibiotics grew in popularity after World War II, interest in the fat-soluble vitamins waned and cod liver oil use began its steady decline (Figure 1).9
The emergence of molecular biology in the late twentieth century provided new evidence for synergy. Vitamins A and D both make independent contributions to immune function by binding to their respective receptors and thereby directing cellular processes in favor of healthful immune responses, but studies in isolated cells suggest that vitamin D may only be able to activate its receptor with the direct cooperation of vitamin A.11,12 We now know that vitamins A and D also cooperate together to regulate the production of certain vitamin K-dependent proteins. Once vitamin K activates these proteins, they help mineralize bones and teeth, support adequate growth, and protect arteries and other soft tissues from abnormal calcification, and protect against cell death.1
As described below, the synergistic action of the fat-soluble trio depends on support from other nutrients like magnesium, zinc, fat and carbohydrate, as well as important metabolic factors such as carbon dioxide and thyroid hormone.
Magnesium and the Fat -Soluble Vitamins
Magnesium contributes to more than three hundred specific chemical reactions that occur within our bodies, including every reaction that depends on ATP, the universal energy currency of our cells.13 Magnesium also activates the enzyme that makes copies of DNA, as well as the enzyme that makes RNA, which is responsible for translating the codes contained within our genes into the production of every protein within our body. This process of translating the DNA code in order to produce proteins is called “gene expression.”
Vitamins A and D carry out most of their functions by regulating gene expression, which means they rely directly on magnesium to carry out these functions. They also rely indirectly on magnesium because our cells can only produce their receptors and all the proteins with which they interact with the assistance of this critical mineral.
The well-studied interaction of magnesium with vitamin D and calcium provides an illustrative example.13 Magnesium is required for both steps in the activation of vitamin D to calcitriol, the form of vitamin D that regulates gene expression and stimulates calcium absorption. Even fully activated vitamin D (calcitriol), however, is useless in the absence of magnesium. Humans who are deficient in magnesium have low blood levels of both calcitriol and calcium, but treating them with calcitriol does nothing to restore calcium levels to normal. The only way to normalize calcium levels in these subjects is to provide them with sufficient magnesium. Magnesium also supports the cellular pumps that keep most calcium out of our soft tissue cells and make it available for the extracellular matrix of bones and teeth.
Altogether, these findings suggest that vitamins A and D can only fulfill their functions in the presence of adequate magnesium. As can be seen in Table 1, magnesium is abundant in many whole grains, nuts, seeds, legumes, and vegetables, some fruit, and some seafood. It is less abundant in meat, by contrast, and almost entirely absent from refined grains and sugar.14 Thus, we would expect a well-rounded diet devoid of “the displacing foods of modern commerce” to be rich in magnesium and thereby support the synergistic action of the fat-soluble vitamins.
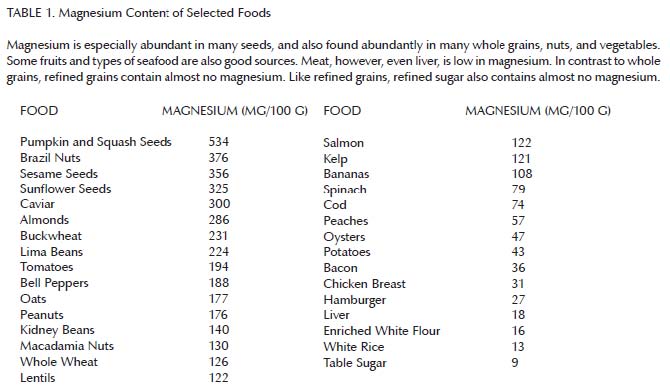{kind=link}
Zinc and the Fat -Soluble Vitamins
As with magnesium, the fat-soluble trio can only support health if our diets contain adequate zinc. The interaction between vitamin A and zinc is particularly well studied.15 Vitamin A supports the intestinal absorption of zinc, possibly by increasing the production of a binding protein in the intestines. Zinc, in turn, supports the formation of vesicles involved in transporting vitamin A and the other the fat-soluble vitamins across the intestinal wall.
Zinc is an essential structural component of many vitamin A-related proteins, including the primary protein that transports vitamin A through the blood, the enzyme that carries out the first step in the activation of vitamin A to retinoic acid, and the nuclear receptor that binds to retinoic acid and allows it to regulate gene expression.
Numerous studies have demonstrated the interaction between zinc and vitamin A in humans. For example, in humans with marginal zinc status, zinc supplementation supports vitamin A’s role in visual function16 (see sidebar below) and eye development (Figure 2).17
Although less well studied, zinc also interacts with vitamin D. Vitamin D and zinc most likely promote each other’s intestinal absorption.15,18 In rats, dietary zinc supports the production of the vitamin D receptor.19 Once the receptor is formed, zinc provides it with essential structural support. Although in the absence of this structural support the receptor still binds to vitamin D, the structural support is needed to allow this vitamin-receptor complex to bind to DNA.20 Studies with isolated cells illustrate the importance of this interaction: adding zinc to these cells increases the rate at which vitamin D activates the expression of genes.21
Altogether, these results suggest that vitamins A and D can only fulfill their functions in the presence of adequate zinc. As can be seen in Table 2, zinc is found most abundantly in oysters, beef, and cheese.22 Chicken and eggs have considerably less zinc than beef, while milk and most plant foods contain very little. We absorb zinc about five times more effectively from animal products than from plant products. Thus, we should expect a diet that contains liberal quantities of zinc-rich animal products to support the synergistic action of the fat-soluble vitamins.
Fat , Carbs , Thyroid and Carbon Dioxide
In order to absorb fat-soluble vitamins from our food, we need to eat fat. Human studies show that both the amount and type of fat are important. For example, one study showed that absorption of beta-carotene from a salad with no added fat was close to zero. The addition of a lowfat dressing made from canola oil increased absorption, but a high-fat dressing was much more effective.23 Canola oil, however, is far from ideal. Studies in rats show that absorption of carotenoids is much higher with olive oil than with corn oil.24
Similarly, studies in humans show that consuming beta-carotene with beef tallow rather than sunflower oil increases the amount we absorb from 11 to 17 percent.31 The reason for this is unknown, but it may be that oils rich in polyunsaturated fatty acids promote the oxidative destruction of fat-soluble vitamins in the intestines before we are able to absorb them. Thus, the more fat we eat, and the lower those fats are in polyunsaturated fatty acids, the more fat-soluble vitamins we absorb.
While dietary fat is clearly important, there may be a role for dietary carbohydrate as well. Once vitamins A and D stimulate the production of vitamin K-dependent proteins, vitamin K activates those proteins by adding carbon dioxide to them. Once added to a protein, carbon dioxide carries a negative charge and allows the protein to interact with calcium, which carries a positive charge. The greater the supply of carbon dioxide, the better vitamin K can do its job.25 Carbohydrates are rich in carbon and oxygen, and when we break them down for energy we release these elements in our breath as carbon dioxide. Because carbohydrates are richer in oxygen, burning them generates about 30 percent more carbon dioxide per calorie than burning fat, and low-carbohydrate diets have been shown to lower blood levels of carbon dioxide (Figure 3).26 Ideally, we should study this further by determining whether dietary carbohydrate affects the amount of activated vitamin K-dependent proteins in humans.
We also produce more carbon dioxide when we burn more calories, regardless of whether we are burning carbohydrate or fat. Intense exercise more than doubles the amount of carbon dioxide we produce compared to what we produce when at rest.27 Even working at a standing desk rather than a sitting desk increases both calories burned and carbon dioxide generated by about a third (Figure 4).28
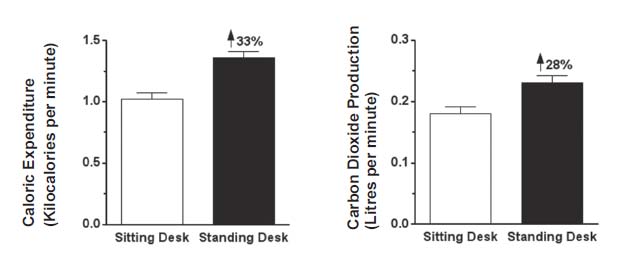{kind=link}
Future studies should directly investigate whether exercise increases the activation of vitamin K-dependent proteins, but it seems reasonable to suggest that part of the reason exercise promotes cardiovascular health may be because it ensures a more abundant supply of carbon dioxide, which vitamin K uses to activate proteins that protect our heart valves and blood vessels from calcification. Thyroid hormone is a key regulator of the metabolic rate and may thus be a major determinant of the carbon dioxide available for activating vitamin K-dependent proteins. Theoretically, thyroid hormone should increase the rate of metabolism and a greater rate of metabolism should produce a proportionally greater supply of carbon dioxide.
Thyroid hormone directly increases the production of vitamin K-dependent proteins and protects blood vessels from calcification in rats.29 The reason for this relationship is unclear. We could speculate, however, that our bodies in their infinite wisdom use thyroid hormone to tie the production of vitamin K-dependent proteins to the production the carbon dioxide needed to activate them.
The Big Picture
It is clearly time to move beyond viewing each vitamin in isolation. The fat-soluble vitamins not only synergize with each other, but cooperate with many other nutrients and metabolic factors such as magnesium, zinc, fat, carbohydrate, carbon dioxide and thyroid hormone.
This paradigm has two important implications. At the level of scientific research, a study about one vitamin can easily come to false conclusions unless it takes into account its interactions with all the others. We should reverently and humbly bow before the complexity of these interactions, realizing how little we know and recognizing that we are always learning. At the level of personal health, these interactions emphasize the need to consume a well-rounded, nutrient-dense diet. Supplementation with an individual vitamin runs the risk of throwing it out of balance with its synergistic partners. The fat-soluble vitamins work most safely and effectively when we obtain them from natural foods within the context of a diet rich in all their synergistic partners.
REFERENCES
1. Masterjohn C. On the Trail of the Elusive X Factor: A Sixty-Two-Year-Old Mystery Finally Solved. Wise Traditions. 2007;8(1).
2. Green HN, Mellanby E. Vitamin A as an Anti-Infective Agent. BMJ. 1928;2:691-6.
3. Semba RD. Vitamin A as “anti-infective” therapy, 1920-1940. J Nutr. 1999;129(4):783-91.
4. Champe PC, Harvey RA, Ferrier DR. Lippincott’s Illustrated Reviews: 3rd Edition. Lippincott Williams & Wilkins. Baltimore, Philadelphia. 2005; p. 387
5. Masterjohn C. Vitamin A on Trial: Does Vitamin A Cause Osteoporosis? Wise Traditions. 2006;7(1).
6. Holmes AD, Pigott MG, Sawyer WA, Comstock L. Vitamins Aid Reduction of Lost Time in Industry. Indust Eng Chem. 1932;24:1058-60.
7. Hospital for Consumption and Diseases of the Chest. The First Medical Report of the Hospital for Consumption and Diseases of the Chest, presented to the Committee of Management by the Physicians of the Institution. J Churchill, 1849, pp. 31-41.
8. Ellison JB. Intensive Vitamin Therapy in Measles. BMJ. 1932;2:708-11.
9. USDA Statistical Bulletin No. 376, U.S. Fats and Oils Statistics, 1909-65: Oilseeds, Oils and Meals, Animal Fats and Oils, Food Fats, Nonfood Fats.
10. Spiesman IG. Massive Doses of Vitamins A and D in the Prevention of the Common Cold. Arch Otolaryngol Head Neck Surg. 1941;34(4): 787-791.
11. Sanchez-Martinez R, Castillo A, Steinmeyer A, Aranda A. The retinoid X receptor ligand restores defective signaling by the vitamin D receptor. EMBO Rep. 2006;7(10):1030-4.
12. Sánchez-Martínez R, Zambrano A, Castillo AI, Aranda A. Vitamin D-dependent recruitment of corepressors to vitamin D/retinoid X receptor heterodimers. Mol Cell Biol. 2008;28(11):3817-29.
13. Rude RK & Shils ME. Magnesium. In Modern Nutrition in Health and Disease(eds M.E. Shills et al.) 223-247 (2006).
14. SelfNutritionData: know what you eat., http://nutritiondata.self.com.
15. Christian P, West KP. Interactions between zinc and vitamin A: an update. Am J Clin Nutr. 1998;68(supple):435S-41S.
16. Russell RM. The vitamin A spectrum: from deficiency to toxicity. Am J Clin Nutr. 2000;71:878-84.
17. Udomkesmalee E, Dhanamitta S, Sirisinha S, Charoenkiatkul S, Tuntipopipat S, Banjong O, Rojroongwasinkul N, Kramer TR, Smith JC. Effect of vitamin A and zinc supplementation on the nutriture of children in Northeast Thailand. Am J Clin Nutr. 1992;56:50-7.
18. Fleet JC, Turnbull AJ, Bourcier M, Wood RJ. Vitamin D-sensitive and quinacrine-sensitive zinc transport in human intestinal cell line Caco-2. Am J Physiol. 1993;264(6 Pt 1):G1037-45.
19. Yu XD, Yan CH, Yu XG, Gao Y, Xu J, Shen XM. Effect of zinc deficiency on the protein expression of vitamin D receptor and calcium binding protein in growth-stage rats duodenal mucosa. Zhonghua Er Ke Za Zhi. 2006;44(1):11-4.
20. Craig TA, Benson LM, Naylor S, Dumar R. Modulation effects of zinc on the formation of vitamin D receptor and retinoid X receptor alpha-DNA transcription complexes: analysis by microelectrospray mass spectrometry. Rapid Commun Mass Spectrom. 2001;15(12):1011-6. 21. Lutz W, Burritt MF, Nixon DE, Kao PC, Kumar R. Zinc Increases the Activity of Vitamin D-Dependent Promoters in Osteoblasts. Biochem Biophys Res Commun. 2000;271(1):1-7.
22. Gropper SS, Smith JL, Groff JL. Advanced Nutrition and Human Metabolism: Fourth Edition. Boston, MA: Wadsworth (2004).
23. Brown MJ Ferruzzi MG, Nguyen ML, Cooper DA, Eldridge AL, Schwartz SJ, White WS. Carotenoid bioavailability is higher from salads ingested with full-fat than with fat-reduced salad dressings as measured with electrochemical detection. Am J Clin Nutr. 2004;80(2):396-403.
24. Clark RM, Yao L, She L, Furr HC. A comparison of lycopene and astaxanthin absorption from corn oil and olive oil emulsions. Lipids. 2000;35(7):803-6.
25. Rishavy MA, Hallgren KW, Berkner KL. The vitamin K-dependent carboxylase generates [gamma]-carboxylated glutamates by using CO2 to facilitate glutamate deprotonation in a concerted mechanism that drives catalysis. J Biol Chem. 2011;286(52):44821-32.
26. al-Saady NM, Blackmore CM, Bennett ED. High fat, low carbohydrate, enteral feeding lowers PaCO2 and reduces the period of ventilation in artificially ventilated patients. Intensive Care Med. 1989;15(5):290-5.
27. Chamari K, Ahmaidi S, Fabre C, Ramonatxo M, Prefault C. Pulmonary gas exchange and ventilatory responses to brief intense intermittent exercise in young trained and untrained adults. Eur J Appl Physiol Occup Physiol. 1995;70(5):442-50.
28. Reiff C, Marlatt K, Dengel DR. Difference in caloric expenditure in sitting versus standing desks. J Phys Ac Health. 2012;9(7):1009-11.
29. Sato Y, Nakamura R, Satoh M, Fujishita K, Mori S, Ishida S, Yamaguchi T, Inoue K, Nagao T, Ohno Y. Thyroid hormone targets matrix Gla protein gene associated with vascular smooth muscle cell calcification. Circ Res. 2005;97(6):550-7.
30. Oregon State University Linus Pauling Institute Micronutrient Information Center. Vitamin A. http://lpi.oregonstate.edu/infocenter/vitamins/vitaminA/index.html Published December, 2003. Accessed November 23, 2012.
31. Hu X, Jandacek RJ, White WS. Intestinal absorption of [beta]-carotene ingested with a meal rich in sunflower oil or beef tallow: postprandial appearance in triacylglycerol-rich lipoproteins in women. Am J Clin Nutr. 2000;71:1170-80.
Zinc and the Dark Adaptation Test for Vitamin A Deficiency
The role of vitamin A in vision is unusual. This vitamin carries out most of its known actions by regulating the expression of specific sets of genes. Vitamin A regulates gene expression only after being activated in a two-step process from retinol to retinal, and finally to retinoic acid. Vitamin A supports vision, however, in its semi-activated form as retinal. Retinal binds to a protein known as opsin, forming a vitamin-protein complex known as rhodopsin. Each photon of light that enters our eye and collides with rhodopsin causes the retinal to change shape and release itself from the complex. This event then translates into an electrical impulse that our optic nerve transmits to our brain. The brain synthesizes myriad such electrical impulses at every moment and interprets them as vision.30
While the function of opsin is to help generate visual images by binding and releasing vitamin A, opsin can only maintain its proper shape and function when it is bound to zinc. In addition, zinc supports the conversion of retinol to retinal, the form of vitamin A that binds to opsin. We could predict, then, that vitamin A would only be able to support vision in the presence of adequate zinc. This can be studied by determining dark adaptation thresholds, which determine the dimmest spots of light we are able to see after having spent a period of time in the dark to maximize our visual sensitivity. When vitamin A is insufficient, we lose the ability to see the dimmer spots of light.
Robert Russell of Tufts University studied ten patients with deficient blood levels of vitamin A who also failed the dark-adaptation test. Eight of them achieved normal dark-adaptation thresholds after supplementing with 10,000 international units of vitamin A for two to four weeks. Two of them, however, had deficient blood levels of zinc. Vitamin A supplementation alone failed to normalize their visual function, but adding 220 milligrams per day of zinc to the regimen for two weeks brought it back to normal.16 These results show that vitamin A can only support healthy vision with the direct assistance of zinc.
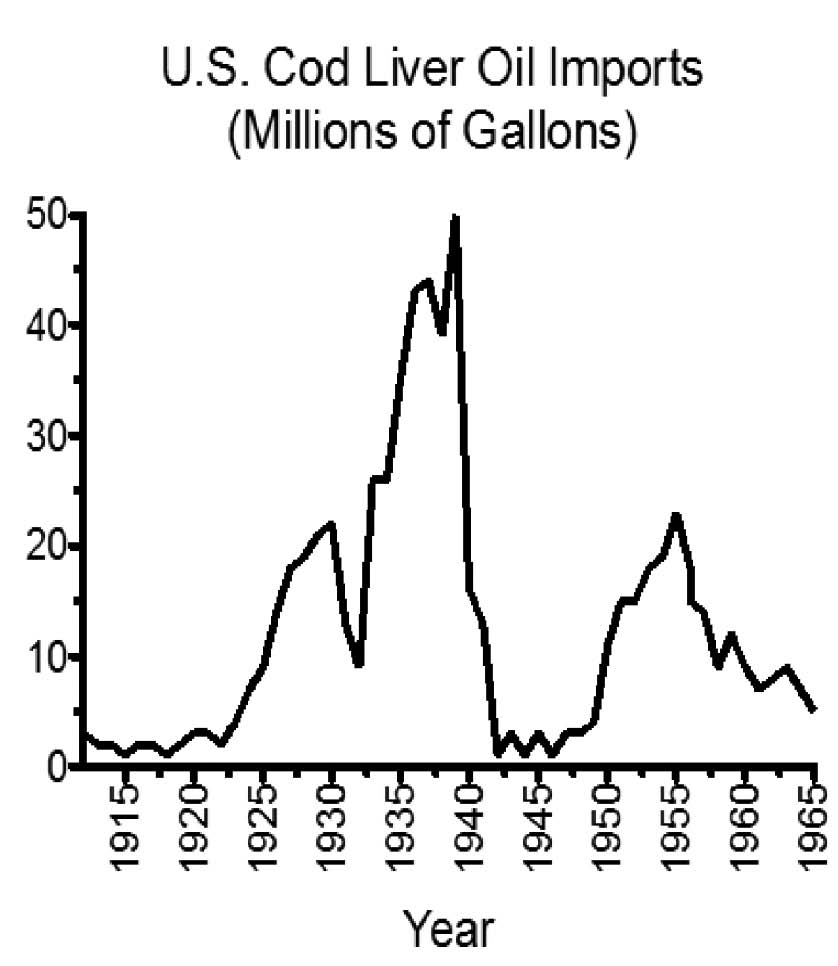
FIGURE 1:
Cod liver oil imports to the U.S., 1912-1965. Adapted from the data in reference 9.
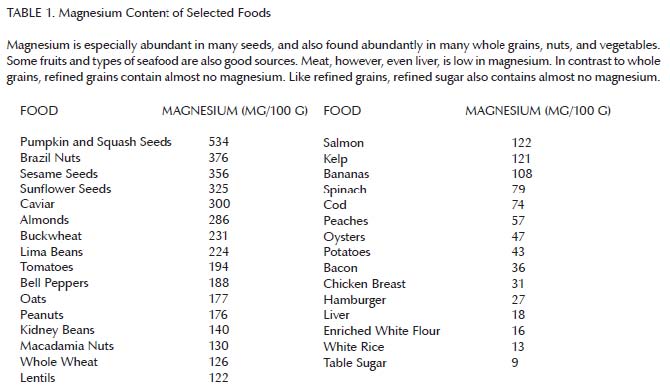
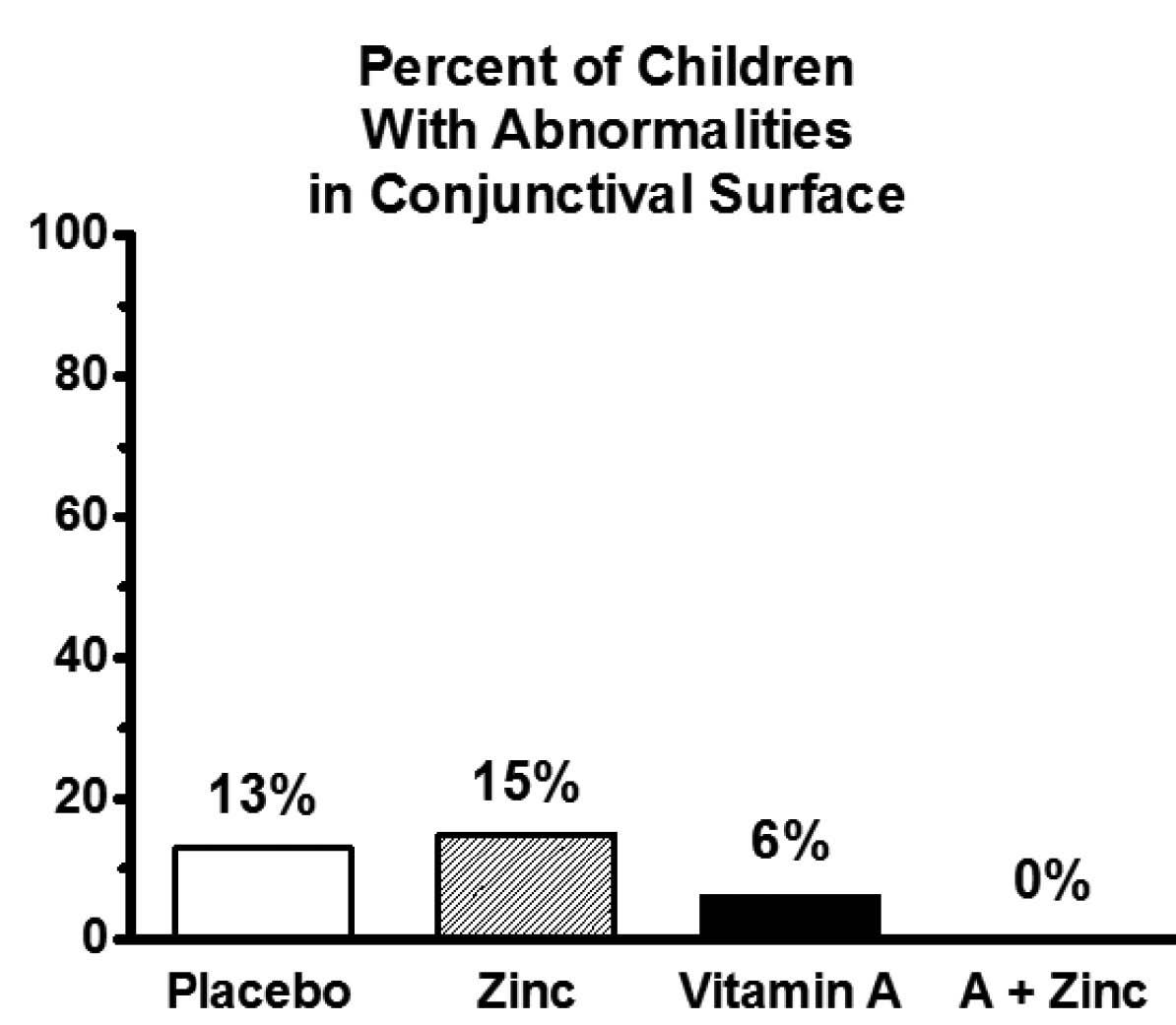
Figure 2 Thirty-three Thai children ages six to thirteen with marginal zinc and vitamin A status were randomized in double-blind fashion to receive a placebo, 25 mg/day zinc, 1300 international units/day vitamin A, or a combination of the two supplements for six months. The figure shows the proportion of children with abnormalities in the conjunctiva of the eye, a characteristic sign of vitamin A deficiency. The synergistic interaction between zinc and vitamin A is statistically significant (P<0.02). The development of the surface of the eye and many other tissues is regulated by retinoic acid, the fully activated form of vitamin A. These results show that zinc supports the role of vitamin A in tissue development, probably by supporting the activity of enzymes involved in activating vitamin A to retinoic acid and by allowing the nuclear receptors for retinoic acid to bind to DNA and regulate gene expression. Adapted from reference 17.
TABLE 2 Zinc Content of Selected Foods
Zinc is found most abundantly in oysters, beef, and cheese. Although not reflected in the table, zinc is generally absorbed from animal products about five times more effectively than plant products.
Oysters
Ground Beef
Liver
Cheese
Chicken
Eggs
Legumes
Milk
Grains and Cereals
Vegetables
Fruit
17-91
3.9-4.1
3.1-3.9
2.8-3.2
1.0-2.0
1.1
0.6-1.0
0.4
0.3-1.0
0.1-0.7
<0.1
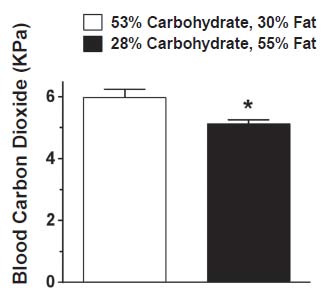
Figure 3: Low-Carbohydrate Diets Reduce Blood Levels of Carbon Dioxide
Investigators fed twenty adult patients requiring artificial ventilation a standard or low-carb, high-fat diet through a feeding pump. The purpose of the study was to determine whether a low-carbohydrate diet could be used to reduce the need for a ventilator. Carbohydrates generate more carbon dioxide than fat, and we eliminate excess carbon dioxide by breathing faster. The study showed that patients on a low-carbohydrate diet generated less carbon dioxide, breathed more slowly, and required less time on a ventilator. Nevertheless, for healthy individuals who do not require artificial ventilation, dietary carbohydrates may support the activity of vitamin K, which activates certain proteins by adding carbon dioxide to them. Adapted from reference 26.
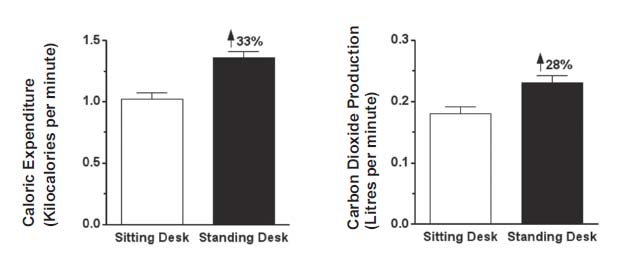
Figure 4: Twenty healthy young adult men and women performed crossword puzzles and word finds for forty-five minutes either at a traditional sitting desk or at a standing desk. Adapted from reference 28.
This article appeared in Wise Traditions in Food, Farming and the Healing Arts, the quarterly journal of the Weston A. Price Foundation, Winter 2012.
Zinc
According to the USDA, nuts/seeds are also a great source of zinc even if they are not as highly absorbed as that from animal products. Sprouting and lightly roasting nuts/seeds helps reduce the phytates. Pumpkin seeds especially, are a great source of zinc at 7.81 mg/100g.
No nut or seed comes anywhere close to the zinc supplied by oysters.
Zinc is found most abundantly in oysters, beef, and cheese. Although not reflected in the table, zinc is generally absorbed from animal products about five times more effectively than plant products.
This is the best paper I’ve read yet on the synergistic relationships of minerals and vitamins. It might be added too, that because minerals also have antagonistic relationships, when you have excesses in one (say copper) its common to have a deficiency in it’s antagonist (zinc). Multi-vitamins and/or multi-minerals are a VERY bad idea because most people have some level of imbalances these days, and taking a multi’s can contribute to both excesses and deficiencies.
Thanks for a great piece of writing.
Interesting research which contradicts what some sources say in terms of the synergies between Vitamin A and D. Most sources indicate they are antagonists, yet I always found that odd since they are found together in nature (cod livers as an example) and seem to perform important functions in conjunction with one another.
Antagonist is a term used because modern people are caught up in Hegelian dialectictics.
Cooperative systems are labeled antagonists, like the operation of male erection and ejaculation vice the sympathetic and parasympathetic nervous system. No, neyat no. Is cyclical pattern of mutual dependance.
Regardless. The dark purpose of such antagonistic semantics is mass confusion. Conventional science editors jump at such confounding rhetoric.
EXCELLENT WORK
We also have vitamin k which acts by supporting blood clotting
Which vegetable K is K1, that the statin drug pushers seek to exterminate – along with the animal fats version, K2 (way too healthy to be healthy for drug sales), implicated by adsociation.
Begin at the beginning:
Throughout the twentieth century, we viewed the fat-soluble vitamins in isolation from one another. Researchers relegated vitamin K to blood coagulation, and ignored vitamin K2 entirely because it is present in such small quantities in the diet. Vitamins A and D each fell in and out of favor, the popularity of one always rising at the expense of the other. Ignoring the forest to study one tree at a time, the concept of synergy eluded us.
Parabéns!
O mundo está a mudar, caindo barreiras da ciência e crenças muito rapidamente. Na difusão da evolução, sois um pilar na melhor visão esclarecida.
Faz todo o sentido como o vedes e, difundindo, ajudais a construi-lo mais verdadeiro.
Obrigado.
Natural Foods for the win! Good Article
Can I use encapsulated food supplements to meet nutritional needs?